Статьи
Физиология мышц
14 октября 2012
Комментариев: -
Поделиться на:

В предыдущих темах мы с вами пытались разобраться в терминах и определили, что такое тренировка с точки зрения физиологии и с точки зрения педагогического процесса. Но вот практический вопрос.
После некоторого периода кроссовой подготовки вы переходите на стадион, проводите одну скоростную тренировку и на следующий день испытываете сильную боль в мышцах. Казалось бы длительный кроссовый бег должен был подготовить мышцы. Что по сравнению с ним короткие отрезки, когда даже и устать не успеваешь?
Но мышцы болят, и для того чтобы узнать, что с ними происходит, нужно разобраться, как они работают: начинают ускорение и торможение, придавая движение нашему телу. В этом нам поможет "Физиология мышц" Й. Рюэгг.
Взаимодействие человека с внешней средой не может осуществляться без сокращений его мышц. Масса мышц намного больше, чем других органов; они составляют 40-50% массы тела. Мышцы - это «машины», преобразующие химическую энергию непосредственно в механическую (работу) и в теплоту. Деятельность их, в частности механизм укорочения и генерирования силы, сейчас можно достаточно детально объяснить на молекулярном уровне с использованием физических и химических законов.
1. Молекулярный механизм сокращения
Один грамм ткани скелетной мышцы содержит примерно 100 мг «сократительных белков» - актина (молекулярная масса 42000) и миозина (молекулярная масса 500000). Механизм их взаимодействия во время элементарного акта мышечного сокращения объясняет теория скользящих нитей, разработанная Хаксли и Хансон.
Теория скользящих нитей
Сократительные белки актин и миозин образуют в миофибриллах тонкие и толстые миофиламенты. Они располагаются параллельно друг другу внутри мышечной клетки, как показано на рис. 8 - схеме крошечного участка мышечного волокна человека. На ней изображены также одна из митохондрий (саркосом), лежащая между миофибриллами, и часть системы поперечных и продольных трубочек.
Миофибриллы представляют собой сократимые пучки «нитей» (филаментов) диаметром около I мкм. Перегородки, называемые Z-пластинками, разделяют их на несколько компартментов - саркомеров - длиной примерно по 2,5 мкм.
Структура саркомеров схематически показана на рис. 8. С помощью светового микроскопа в них можно видеть регулярно чередующиеся поперечные светлые и темные полосы. Согласно теории Хаксли и Хансон, такая поперечная полосатость миофибрилл обусловлена особым взаиморасположением актиновых и миозиновых филаментов. Середину каждого саркомера занимают несколько тысяч «толстых» нитей миозина диаметром примерно по 10 нм. На обоих концах саркомера находятся около 2000 «тонких» (толщиной по 5 нм) нитей актина, прикрепленных к Z-пластинкам наподобие щетинок в щетке. Пучок лежащих в определенном порядке миозиновых нитей длиной 1,6 мкм в середине саркомера выглядит в световом микроскопе темной полосой шириной 1,6 мкм; из-за свойства двойного лучепреломления в поляризованном свете (т. е. анизотропии) она называется А-диском. По обе стороны от А-диска находятся изотропные участки, содержащие только тонкие нити и поэтому выглядящие светлыми; эти так называемые I-диски тянутся до Z-пластинок. Именно в результате такого периодического чередования светлых и темных полос в бесчисленных саркомерах миофибриллы сердечной и скелетной мускулатуры выглядят поперечнополосатыми.
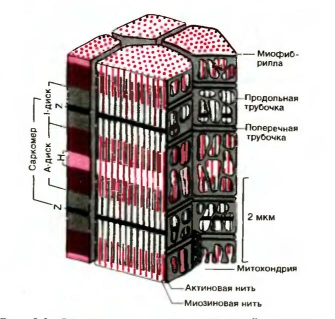
Рис. 8. Схема участка волокна скелетной мышцы человека (по Garamvolgyi)
В покоящейся мышце концы толстых и тонких филаментов обычно лишь слабо перекрываются на границе между А- и I-дисками. Эта зона перекрывания в А-диске выглядит в световом микроскопе гораздо темнее центральной Н-зоны, в которой нет актиновых нитей. На электронных микрофотографиях Н-зоны видна очень тонкая темная М-линия в середине саркомера - сеть опорных белков, по-видимому, удерживающих толстые нити в составе единого пучка.
Укорочение саркомеров. Мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах. Сравнивая структуры саркомера в двух различных функциональных состояниях (рис. 9), можно видеть изменения поперечной исчерченности и взаиморасположения нитей во время сокращения: тонкие актиновые филаменты скользят вдоль толстых миозиновых, двигаясь между ними к середине их пучка и саркомера.
Рис. 9 иллюстрирует основное положение теории скользящих нитей - во время сокращения мышцы сами актиновые и миозиновые нити не укорачиваются. Это объясняет данные световой микроскопии: ширина А-диска (1,6 мкм) всегда остается постоянной, тогда как I-диски и Н-зоны при сокращении сужаются.
Длина нитей не меняется и при растяжении мышцы. Тонкие филаменты попросту вытягиваются из промежутков между толстыми нитями, так что степень перекрывания их пучков уменьшается.
Каким же образом осуществляется «разнонаправленное скольжение» актиновых нитей в двух половинах саркомера?
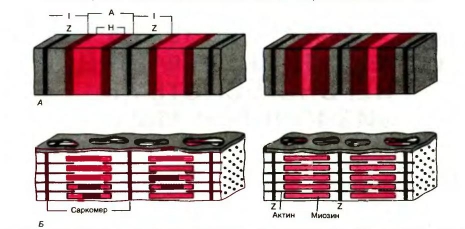
Рис. 9. А. Поперечнополосатая структура миофибрилл: слева - расслабление, справа - сокращение. Б. Организация миозиновых и актиновых нитей в расслабленном и сократившемся саркомере. Обратите внимание на аддитивный характер укорочения последовательно соединенных саркомеров.
Работа поперечных мостиков. Миозиновые нити несут поперечные, отходящие биполярно, как показано на рис. 10, А, выступы длиной около 20 нм с головками примерно из 150 молекул миозина. Во время сокращения каждая головка (поперечный мостик) может связывать миозиновую нить с соседними актиновыми (рис. 10, А). Движение головок создает объединенное усилие, как бы «гребок», продвигающий актиновые нити к середине саркомера. Сама биполярная организация молекул миозина обеспечивает противоположную направленность (стрелки на рис. 10) скольжения актиновых нитей в левой и правой половинах саркомера.
В результате однократного движения поперечных мостиков вдоль актиновой нити саркомер укорачивается только на 2 х 10 нм, т.е. примерно на 1% своей длины. Однако при изотоническом сокращении мышцы лягушки саркомеры за десятую долю секунды укорачиваются на 0,4 мкм, т. е. на 20% длины. Для этого поперечные мостики должны совершить свои гребковые движения за указанный промежуток времени не один, а 20 раз. Только за счет ритмичных отделений и повторных прикреплений миозиновых головок актиновая нить может подтягиваться к середине саркомера, подобно тому как группа людей тянет длинную веревку, перебирая ее руками. Благодаря суммации минимальных укорочений миофибрилл в последовательно расположенных саркомерах мышца лягушки длиной 2 см при изотоническом сокращении за 0,1 с поднимет маленький груз на высоту 0,4 см. Следовательно, когда принцип «вытягивания веревки» реализуется во множестве последовательных саркомеров, повторяющиеся молекулярные движения поперечных мостиков приводят к макроскопическому движению. При расслаблении мышцы миозиновые головки отделяются от актиновых нитей. Поскольку актиновые и миозиновые нити могут легко скользить друг относительно друга, сопротивление расслабленных мышц растяжению очень низкое. Их можно снова растянуть до исходной длины, приложив совсем небольшое усилие. Следовательно, удлинение мышцы во время расслабления носит пассивный характер. Только что описанные движения поперечных мостиков недавно были продемонстрированы на молекулярном уровне методом дифракции рентгеновских лучей (их малоугловое рассеивание сократившейся мышцей с временным разрешением порядка нескольких миллисекунд).
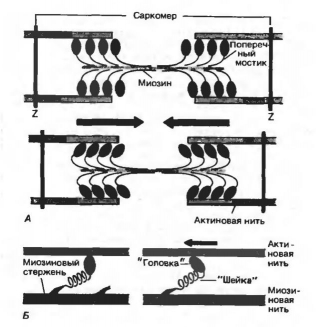
Рис. 10. Функция поперечных мостиков. А. Модель механизма сокращения: миозиновая нить с поперечными мостиками, прикрепленными к соседним актиновым нитям; вверху - до, внизу - после «гребка» мостиков (на самом деле они функционируют асинхронно). Б. Модель механизма генерирования силы поперечными мостиками; слева - до, справа - после «гребка». Поперечные мостики химически соответствуют субфрагменту миозина-«тяжелому меромиозину», который состоит из субфрагментов I (головка) и II (швйка)
Генерирование мышечной силы. Благодаря упругости поперечных мостиков саркомер может развивать силу даже без скольжения нитей относительно друг друга, т. е. в строго изометрических экспериментальных условиях. Рис. 10, Б иллюстрирует такой процесс генерирования изометрической силы. Сначала головка миозиновой молекулы (поперечный мостик) прикрепляется к актиновой нити под прямым углом. Затем она наклоняется под углом примерно 45°, возможно, благодаря притяжению между соседними точками прикрепления на ней и на актиновой нити. При этом головка действует как миниатюрный рычаг, приводя внутреннюю упругую структуру поперечного мостика (видимо, «шейки» между головкой и миозиновой нитью) в напряженное состояние. Возникающее в результате упругое растяжение достигает лишь около 10 нм. Упругое натяжение, создаваемое индивидуальным поперечным мостиком, так слабо, что для развития мышечной силы, равной 1 мН, нужно объединить усилия по крайней мере миллиарда таких соединенных параллельно мостиков. Они будут тянуть соседние актиновые нити, как команда игроков тянет канат.
Даже при изометрическом сокращении поперечные мостики не находятся в непрерывно напряженном состоянии (это наблюдается только при трупном окоченении). На самом деле каждая миозиновая головка уже через сотые или десятые доли секунды отделяется от актиновой нити; однако через такое же короткое время следует новое прикрепление к ней. Несмотря на ритмичное чередование прикреплений и отделений с частотой порядка 5-50 Гц, сила, развиваемая мышцей в физиологических условиях, остается неизменной (исключение-летательные мышцы насекомых), так как статистически в каждый момент времени в прикрепленном, обусловливающем напряжение, состоянии находится одно и то же количество мостиков.
Изометрическое теплообразование. Мышца, поддерживающая определенное сократительное напряжение в изометрических условиях, отличается от сокращающейся изотонически тем, что не выполняет внешней работы (произведение силы на расстояние равно нулю). Однако в каждом цикле прикрепления-отделения поперечных мостиков совершается внутренняя работа по растяжению их упругих структур, которая преобразуется в тепло в момент отсоединения миозиновых головок. Теплота изометрического сокращения («изометрическая работа») за единицу времени возрастает с увеличением количества функционирующих поперечных мостиков и частоты «гребков», требующих расхода АТФ.
Теперь можно представить, о каких микроповреждениях пишет профессор Виктор Николаевич Селуянов во второй части статьи "Сердце - не машина..." В следствии этих микротравм происходит набухание клетки (мышечного волокна - МВ), о чем и сигнализируют нервные окончания, которые находятся на мембране клетки, вызывая болезненные ощущения.
Таким обрязом, изучая строение МВ, его работу и энергообеспечение, можно узнать, что необходимо спортсмену кроме тренировок. Это конечно же соблюдение режима сна и питания, активная повседневная деятельность, положительный настрой и многое другое.
продолжение следует
© 2011 - 2024 Tula Track and field.
Лёгкая атлетика в Туле
При использовании материалов сайта ссылка обязательна.
Сделано в