������
���������� ���� (�����������)
07 ������ 2012
������������: -
���������� ��:

���� �� ����������� ��������� ������ ����� ���� ������, �� � ��� �������� ������ ������ - ��� ���������� ��������� ���������� �������, ������ ��� �� �������������, �� ���������� �� ��������� ����? � ��� ��� ���������� ��������� � ����������� - ��������? ��� ��������� �� ��������� ���������� �������� � ��� ��� ������?����� ������ �������� ��������� �������� ���������� �� ������ ������������� ����������� ��������������� ���������� ��� ���� ������� ������, �� ������� ��� ����������� ������������� �������, ��� ���������� �������� ����� ���� ����, �������� �� �������������� ����� ��� �������������� ������ �� ���������� �����. � ������ ���������� �� ���� �����.
��� ��������� � �� ����, ����� � ������� �� ���������� ������� ������� � ������� ���������� ����� �� ������ ������� �������� ������� ������ �������� ��������� ���������. �� �������, ��� �� ������ �� ��� ��� ������ � ���� ������ ������ ��������. ����� ������ ������, � ����� �� ������, ��� �� ����� �� ���������� ��� �������������� ���-�� �������� � ��������� ���������� � ������ �����-��������.
�������������� ���������� ������� � ������������
����� ������� ����� ����������� ���������� ������� � ������������? ��������, ��� ��������� ������ ����������� ������������ ������������ ����.
���-���������������� �������� ������� ��� ����������. �������������� ������ ������ �� �������� �������� � ��� ���, ��� ���� ������������������ ��������������� ����������� ��� �� ��� � ������� �� ����� ���������� �����. ��� ������ �������������� ������� ������� � ���, �������� �������� � ���������� ����������� ��������� � ������ ��������������, �� ������������ ���� ������� ���������������; ��� ������ ������ ��� ������������ ��������������� �������� ��������-���. �������������� ������� ������� �������� ��������������� � ��������� ��������, ��� ��� ����� �� ����������� ���� ������� ������ (����. 2). �������� ����������� ��� �� ����� ���������� ����� �������� ������ � �������� ������� ��������� ��� ���������������� ��������������� �����. ������������� ����� �������, ������ ������������ ������ ������ �� ��������� ���������� ��������� �������� �������������� ����������, �������� � ������� ���� 2,6 ������ ��� �� 1 � ����� �����, ����� ��� ����������� �����, �� �������������� ����������,- 2,9 ������. ������ ���������������� ��� ���������� ������������� ���������� (0,3 ������) ��������� ������� - ��� � �������. ����� �������, ����������� 0,3 ������ ��� ���������� ������� ��� �������������� ���������� � ��������� �����.
������� 2. ������ � �������� ��������� ������� � ��������� ����� ��������
| ��������� ������� | ����������, ������/� ����� | �������, ������ ������� |
| ����������������� (���) | 5 | ��� -> ��� + � |
| ������������� (��) | 11 | �� + ��� <=> ��� + � |
| ������� (�������� � ������� ���������) | 84 | ���������� ����������� ����� ������� �� ������� (��������) �������� ����������� ����� ������� �� C02 � H2O |
| ������������ | 10 | ��������� �� �02 � �20 |
��� - ����������������, � - �������, P - �������������� ������.
��� �������������� ������������ � �� ���� ����� ������������� ������������ � ����� � ������� ������� �������� - ������ �������, ������ ���� ������� ������������ �������. ����� � ������, ��� ���������� ����, ������������ ����� �������� ���������, ����� ����������� � ������������ ����������, � ���-������������ �������� � ����� (���������� ���������� ������ ������ �������������������), ������� ��� ����� ��������������� ���������������. ������ � �������� ������� �������� ������������ �������������� ���� ������ � ������� (�������������� ����), ��������� ����������� ��� ��, ��� ����� �����, ��������� ��� (������ ���!) � �������� ��������� �������. ��� ������������ ���������������� ������� ��� � �������� ����������.
����������� ��� �� ����� ����������. ������ ��������, ��� ���������� �������, ����������������� � �������, ���� �������� ������������� �������� ������ ��� ����������� ���. ������ ������� ������������ ������� � ����������� Mg2+. �������������, ��� ��������������� ������ ������� �����, �.�. � ����������� Mg2+, ��� ������������, ����������� ��� � ������, ������ � ������ ������������ ������� ������� � ������������� ����� - ������. (� ���������� ������ ������������ ��� �� ��������������, � ��������� �� ��������� ������ �������������� ����� ������� �, ����� �������, ���������� ����������� ���.) � ������ ����� ������������-��������� ����������� ������� ��� ������������ ������ ���� ��� (��������, ���� �������� �� ������ ������). �������������, ��� ������ �������� ��������� � �������� ���������, ��� ���� �������� ����������� ��� � ����, ����������� ������; ������, ��� �������� (������������� �����������), ��� �������, ��������������� ����, ����������� ������.
�������� ��������� ���������� ��� ����, ��� ������� �������� ���������� �������, �.�. ��� ������ �������� ��� ��������� � ������� �������. � ���������� ������� ����� ���������� � ������� ������� ������ � ��������� ��� ���������� ���������� ������ ��� (�������), ��� ���������. ������� ��� ����������� ���� ������������ ��������������� ��������� �������� ������� (���� I), ������� �����������, � ��� ������� �������� - ������ �� ������ (���� IIB) ��� ������� (���� II�) �������.
�������� �������� ���. ��������, � ������� �������� ����� ������� - �������� ��� - ������������ ����������� ����������� �������, ���������� ���������. ��-��������, ��� ����������� � ���������� �������� ����� ���������� �������, ����� ������� ��� ���������� ����������������� �������������� ������ - ������ � �������. ����� ����� �� ����� ����� ���������� ������� ���������� �� ������, � ��� ������������ �� ��� � �������. �������� ��������� �������� �� �������� ����� ���������� � �������������� �������, ��� ���������� ��� ������ ������������� ����������� ������� � ������ � ���������� ������������� ���� �������, �� ����� �������� ���������� ������������� ��� � �������. ����� ��� ��������� ����������� ������� � ��� ������ ��������� ����� �������� ��� � ���������� ����� ����. ��������� ���������� ���������� ��������, �. �. ����� �� ������������ � ������ � ������������ �� ����, �������������� �������� ����������, �������� ������ ��� ��������� ���, � ������, ��� ��������� ������. ���� ����������� ��� �����������, ������� �� ����� ������������� � ������, ������������� ���������� � ���� �������� ������� ������ �� ���� � ����� �������������. ��� ������ ��������� ���������� ��� � ������� ���������; ����� ��� ��������� ����������� �������, ���������� ������� �������� ��������� �������������� � ��������� ���� (���� �� ���������� �������). ��� ���� ���� ������ � ������� ������ ��������� ���� � ������, � ����� ��������� � ��������� �������� ���������� (rigor mortis). ������ �������, �������������� ����������, ���������� � ������������ (����. 3), ������� �� �������� �������������� �������������� ������.
������� 3. ������� ��� �� �������������� ��������� �������� ������� � �� �������������� �����-������
| ���: | ��������� ��������� ������� | ���������� ������� ������� | ������ |
| ����������� | ���������� | ����������� � ������ | - |
| ������������, �� �� ������������ | ������������ | �������� �� ������ | ������������1) |
| ������������, ������������ ������� | ���������� | ����������� ������������ � ��������� | �������2) |
1) ��� ������������ ��2+ < 10-7 M.
2) ��� ������������ ��2+ �������� 10-6 - 10-5 M.
����� �������� ���� ��� � ���������� � ������������, ����� � ��. ������� ������� �� �������� ������� ���� ���������� ��� (��������, ����� ��������������� ������ ��������� ���������, ��� ������ �������� ����������� ��� ���). ����� ������� �������, �� ��� ���������� � ������� ��� ����� ����������� �������� � ������������. ������, ���� ���������� ������ ���������, ��� ��������� �������������� �, ������� ���������� ���� ������������� �������������� �����, ����� ����������� ������ ��� ��������� ������. ��������� ���������� ���������� ������ ����� �������� ������������ ���� ���������� ��������.
2. ��������� ��������� ����������
������ ����� ������������ ��� ����������� ����������� �������� �� ������������� ������������; � ���������� �������� ����������� ����� ������-�������� ������� ������������ �������� ���������� �������� (�������� ����������). �������� � ������ ���������� �������� �������, �� ������ � ����������������� ��������. ��������, ��� ����������� ������������� ����� ������� ��������� ������������� ��������� ������������� ����� 1 �� �� ��������� ������� �� ����� ����������� �������� ����� 1-2 �� �� ��������� �������� 2 �/� ����� ���������������� ��������� ��������, � ��� ����� ��������� ����������� ��� ����������. ����� �������, ���������� ���������� ����������� ��������, �. �. ������������ �������� �������.
������������������� ����������
�������� ������� � ���������� �� ������������ ��������� �������� � ������������ � ������� ������ (������������������� ����������) �������� � ���� ��������� ���������������� ��������� (����. 4), �������� ���� � ������� ������ ���� ��2+.
����������� � �������� �������� ��2+. �������� ��2+ � �������� ������� �������� �� ����������. ��������� ����� ������� ������� ������ �������� ��� ������������ ������� ����������� ��2+ �� �����������, ��� �� �� ������� ����� �������� ��� ���������� ������������� ��������� ��������. ��� ����� �� ���� ��������� (����������) �����������, ���� ������������ ������������, ���� ���������� ������������� ���� ��������������� ����������. ����� �������� ���������� (��������������) �������� ������� ����������� ������ ��� ���������� � �������, ���������� ��� � �� ������� ���� 10-6 � ��������������� ������� ��� ��������� ������.
� ���� �������� ���������� ������� ���������� ����� ����� �� ���� ����������� ����������� ��� ���������� ����������������� � ���������� ������. ���� ������������ ������ ��2+ ������� �� ����� (��������, ������� ����������� ��� ��������), ����������� �������������, ��������� �������������� ����� ����������� ��������� � ������� ���������������, � ������, ����������� ���������� ������ (��. ����. 3). ����� ������ ��������� ������� � � ������ � ��������� ���������� ���������. �� ����������� ��������� ������������ ��2+ �� 10-7 �� 10-5 � ��� ��������� ����������� ����������� ���� ���������� � ���������� ������, ������ ��� ���� ��������� ��������� ��������� ��� ������������ ��2+ 10-6 - 10-5 �.
������� 4. ����� ������������� ����������
1. ���������� ��������� �������
2. ��������� �������� (����������� ��������)
3. ������������������� ����������
�. ���������� ����������� �� �-�������
�. ������������� ��2+ �� ���������� ������� (���. 12)
�. �������� ��2+ �� ����������� (���. 11)
4. ���������� ����������: ����������� ���������� ���������� ��������
�������� ��������� ������ ������� ��������� ������� ����� ������, ���������� ��������� ��������� ����� (���. 11). ������ ����� �������� ������ ����� 1 ��� � �������� 5-7 �� ������� �� ���� ����������� ���� ������ ������ ������� ��������� ������ �������� 5 ��. ������� ��������� ���������, ���� ����� ��� ���� ��� � �������� �� � ���� ������� �� 14 ����� � ������ ����� (���. 11, �). ����� ���������� ���������� �������� �� 40 �� ��������� ������� ����� ����������� �������� ���������, � � �������� ����� ����� ��������� ����� ���� ������������. ������������ � ������� �������������������� ������� (����������� ������������� ���������) ��������, ��� � ���������� ��2+ , �.�. ��� ������������� ��������� ����������, ������� �������� ������������ ������������� ���, ��� ��������� ������������ ���������� ���������� �������� � ��������� �����. � ��������, ��� �������� ��2+ �������� ������������ ������ ���������� � ������� ����� ��������� ��������� ������, �������� ������� ������������ ��� ���������� ��������. � ���������� �� ������������� � ��������� ����� (���. 11, �), ������������ ��� � ����������� �������� ����.
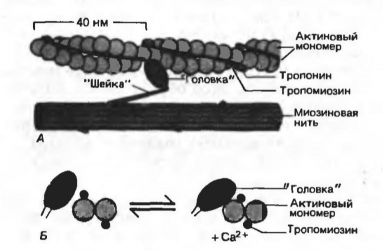
���. 11. �������� ��2+ �� ����� ��������� �����������.
� - ��������� � ���������� ���� �� ���������� ������� �������.
� - ��� �� �� ��� ���������� �������. ����� ��2+ ����������� � ����������, ����������� �������� � ������� ����� ����� ���������� ������, ������� ������� ������������ ���������� ��������
����� �������� ��������� ���������� ��������� ��2+ �� ��������, ������� �������� ��� ����������� ��������������: ��� ���������� � ��2+ ��� �������� ������������� ����� �������, ��� ��� �� ����������� ����������� � ������� ����� ����� ��������� ��������� ���������, �. �. � ��������������� ���������.
�������� � ������������� ����� �������. ������������� ����� �������� ����� 1 ������ ��2+ �� 1 � ����� �����. ���� �� ���� ������� �� ���� ����������� � ������ ��������������� ����������, ����������� ��� ������ �������� ������� ���������� �� � ��������� ������������ ����������.
��������� ��������������� ������ �������� ������� � ������ ������ �� ������ ��������� (��������� ����� �������� - ���. 1; ����� ������� - ���. 12). �� ������ �������� ������������� �������� �������� ������ �������� ���������� � ���� �������� (��������� 50 ��), ���������������� ���������� ��� �������; ��� ������� ���������� �������� ����������� � ������������ ������ � ������ �������� ������ ����������� �� ������ Z-��������� (� �������) ��� � ������� I-������ (� ������ �����������).
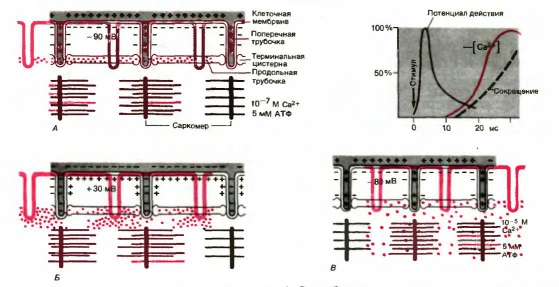
���. 12. ����� �������������������� ����������.
� - ������������� �������� ������� � �������������� ��������� ���������. ������������ ��2+ � ��� ���� 10-7 �.
� - ��������� �������� ������ ���������� �������� ������ � ���������� �������� �� ���������������; ��2+ �������� �������� �� ������������ �������.
� - � ������� ������������ ���������� �������� ��������������� ������������ ��2+ ��������� �������� 10-5 �, � ��������� ���������� �����������. ������ ������, ��������� ������������������ ������� ��� �������-������������ ���������� �� ����������� ������� �� ������ ���������� (���������� ����� ������� ��� 00�)
��������������� ���������� ���������, �.�. ����������� ������������, ����������� ������� ���������� �������� (�������� ������������������� ���������). �������� �� �� ������ (������������ ��������) ��������� � ��������� ������� ���������� ��������, ������� ��� ���������� ������. � ���� ��������� � �������� ��������������� �������. � ������� �� ���������� ������� ���������� �� ���������� � ������������ ������. �������� �������������������� ���������� �������� ���������� �� ������� ��� ���������� �����, ������� ������������ �������� ��������� ��2+ �� ��������� � ���������� ��������, ������ ����� ������� �������� �� 10-7 � ����������������� ������������ ���� ����� � ���������� (�������������) �����.
������������������� ���������� ���������� ����������� ��������������� ���������� �������� �� ��������� ���������� ������� ������ ������. ��� ���� ����������� ������ ��������� � ����� �������, ��������� �� ���������� ������� � � �������� ����� �������� ������������� ��2+ �� ������������ ������� �� ��������������� ��������, ���������� �����������, ��� � ����� � ���������� (���. 12).
��� ��������� �������� ���������� ��������������; ������������ ����� ���������� �������� ��������� ������������ ����� ��2+ ����������� ����������� ������ � ������ �������������������� ����������. �������� ����� ��2+ �� ��������� ���� �� ��� ���, ���� �� ������������ � ��� �� ������ �� �������� 10-7 �. ��� ���� ����������� ���������� ������ ������� � �������������� ����� ������� � ����������� ���������, ������� ���������� �� ��������� ����� (��. ����. 3).
��������� ��� ��� �������: "��������������� ����������� ������ �������" � "��������� �������� ���� � ��������� ��������". �� ���� ��� ���������, ��� ����� �������������� ������� � ��������� ������� ����������, ���������� ��������� ��������.
Mg, Ca, P - ����� ������� �� ������ ��������� � ������� ������� � ��� �������� � ������������� ��������, ���� ������ � �������������� ���. �� ������� ��� ���� ������ - ��� �������� (����� ����� � ������) � ��� � ���� ������� ������� (������������ ����������)
� 2011 - 2024 Tula Track and field.
˸���� �������� � ����
��� ������������� ���������� ����� ������ �����������.
������� �